




É muito comum haver a confusão entre o que são biomas, ecossistemas, florestas? Não?! E a tal Floresta Pluvial Tropical é a Floresta Amazônica ou Atlântica? Pensando nessas questões, traremos hoje um apanhado de maiores para menores áreas, da biosfera para o microclima.
A ecologia é a ciência que estuda a relação dos seres vivos, entre eles, e para com o ambiente. Quanto a isso a pesquisa pode ocorrer em vários níveis: adaptações morfológicas de um animal (organismo); estrutura social de um conjunto desses animais (população), sua dinâmica ou diversidade; como essa população interage com outras (comunidade) pela predação/ competição/ parasitismo...; o impacto de todas essas espécies no ambiente e como transformam o mesmo (ecossistema); situações climáticas que envolvem o mundo todo e podem atingir várias espécies de vários continentes (biosfera).
 A organização entre os seres vivos e o local, vai depender obviamente do clima e se observarmos mapas de temperatura ou ventos, do planeta vemos que há certa zonação. Ventos mais intensos, temperaturas mais altas, maior pluviosidade, etc. ocorrem no equador e vão perdendo sua intensidade conforme seguem aos pólos. Esse conjunto de características favorecem algumas características morfológicas e fisiológicas de seus habitantes, fazendo com que haja também zonação de "tipos" de seres vivos em cada latitude do planeta. O tipo de seres que ocorrem num local, torna-o característico e isso é chamado de ecossistema. Quando ecossistemas ocorrem próximos, é lógico que haja semelhança entre esses. Manguezal e restinga têm por exemplo plantas que tem adaptações para excretar sais, o que não ocorreria em uma vegetação ciliar (que circunda um corpo d'água) no continente. O conjunto de ecossistemas, portanto, que reúne vegetação e clima semelhantes, é chamado de bioma. Para melhor visualização os biomas são arranjados de acordo com a vegetação, simplesmente porque é a estrutura mais fácil de ser observada e que é base de cadeia para os demais organismos. Nesse contexto a maioria são florestas, mas, o conceito persiste para ambientes frios e pouco úmidos.
A organização entre os seres vivos e o local, vai depender obviamente do clima e se observarmos mapas de temperatura ou ventos, do planeta vemos que há certa zonação. Ventos mais intensos, temperaturas mais altas, maior pluviosidade, etc. ocorrem no equador e vão perdendo sua intensidade conforme seguem aos pólos. Esse conjunto de características favorecem algumas características morfológicas e fisiológicas de seus habitantes, fazendo com que haja também zonação de "tipos" de seres vivos em cada latitude do planeta. O tipo de seres que ocorrem num local, torna-o característico e isso é chamado de ecossistema. Quando ecossistemas ocorrem próximos, é lógico que haja semelhança entre esses. Manguezal e restinga têm por exemplo plantas que tem adaptações para excretar sais, o que não ocorreria em uma vegetação ciliar (que circunda um corpo d'água) no continente. O conjunto de ecossistemas, portanto, que reúne vegetação e clima semelhantes, é chamado de bioma. Para melhor visualização os biomas são arranjados de acordo com a vegetação, simplesmente porque é a estrutura mais fácil de ser observada e que é base de cadeia para os demais organismos. Nesse contexto a maioria são florestas, mas, o conceito persiste para ambientes frios e pouco úmidos.

Nas temperaturas mais frias ocorrem Florestas Boreal, ou Taiga, onde há formação de serapilheiras e Tundras, também chamadas de permafrost, porque parte da vegetação está constantemente coberta por uma camada de neve.
Com temperatura moderada há Floresta Sazonal Temperara, ou Decídua, e recebe esse nome porque as folhas de suas arvores entram em senescência rapidamente para conversar o resto da planta e assim ficam amareladas e caem; Floresta Pluvial Temperada, que tem inverno ameno e bastantes chuvas; Desertos que são caracterizados pela baixa precipitação e Bosques que são facilmente reconhecíveis pelo tipo de folha de suas arvores, que são duras e chamadas de esclerofilas.
Em temperaturas mais altas estão: os desertos mais conhecidos, como (os quentes) no bioma Deserto Subtropical; Savanas, que incluem no Brasil Cerrado e Caatinga, são vegetações com folhas decíduas e que sofrem periódicos incêndios e por fim a Floresta Pluvial Tropical, que inclui Atlântica e Amazônica, que é um bioma de vegetação densa, com muitas chuvas e é aqui que há a maior biovidersidade.
Na imagem abaixo pode-se notar a zonação de biomas terrestres no planeta.

No encontro do continente com oceano há alagados e estuários. Alagados são biomas que misturam água doce com salobra no subsolo, com formação de lagos, mas, sem fluxo superficial de água, como marismas e apicuns. Nos Estuários há esse fluxo superficial de água e consequentemente mais nichos com maior riqueza de espécies.
Os Oceanos constituem um único bioma, que se divide em zonas de acordo com a profundidade, que influenciará na ciclagem de nutrientes e fotossíntese, o que interfere na teia trófica. De 0 a 200 metros de profundidade o litoral compreende: supralitoral, região sempre acima da altamar, ou maré alta; mediolitoral que ocorre na zona entremarés alta e baixa, estando oras coberto oras descoberto e infralitoral, que está sempre coberto pela baixamar, ou maré baixa. A maioria dos seres vivos do oceano ocorrem no litoral. De 200 a 1000 metros está a zona batial e de 1000 a 4000 a abissal, onde gradualmente aumentam pressão, diminui-se temperatura e produtividade. Na zona hadal, abaixo de 4000 metros de profundidade, toda fonte de alimento vem de bactérias quimiotróficas ou de restos de outros animais que submergem e esse ambiente segue até o assoalho marinho.

A afirmação de que a diversidade diminui do equador para os pólos não desqualifica os biomas desses últimos quanto ao conhecimento. Na verdade, pensa-se que isso se deve ao fato do fator climático e estabilidade do meio. Mutações e novos organismos surgem a todo momento, porém, como a estabilidade ambiental é menor no equador há mais espaço para mais nichos serem ocupados e então, novas espécies, porém nos pólos a maioria dos nichos já têm seus organismos bem adaptados e que dominam o local, simplesmente porque o mesmo tem poucas variações. Estudos em ambas as situações são válidos, porque evidenciam e embasam linhas de pesquisa diferentes.
Referências:
RICKLEFS, R.E. A Economia da Natureza. 6 ed. Editora Guanabara Koogan: Rio de Janeiro, 2010.
A ecologia é a ciência que estuda a relação dos seres vivos, entre eles, e para com o ambiente. Quanto a isso a pesquisa pode ocorrer em vários níveis: adaptações morfológicas de um animal (organismo); estrutura social de um conjunto desses animais (população), sua dinâmica ou diversidade; como essa população interage com outras (comunidade) pela predação/ competição/ parasitismo...; o impacto de todas essas espécies no ambiente e como transformam o mesmo (ecossistema); situações climáticas que envolvem o mundo todo e podem atingir várias espécies de vários continentes (biosfera).

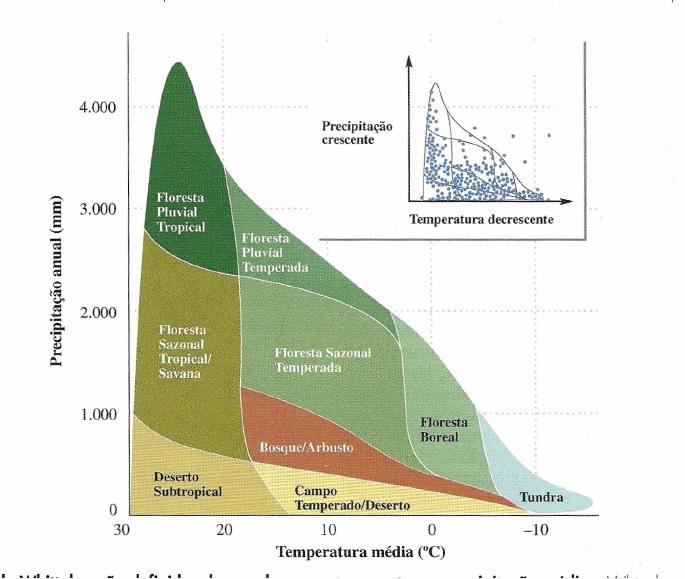
Nas temperaturas mais frias ocorrem Florestas Boreal, ou Taiga, onde há formação de serapilheiras e Tundras, também chamadas de permafrost, porque parte da vegetação está constantemente coberta por uma camada de neve.
Com temperatura moderada há Floresta Sazonal Temperara, ou Decídua, e recebe esse nome porque as folhas de suas arvores entram em senescência rapidamente para conversar o resto da planta e assim ficam amareladas e caem; Floresta Pluvial Temperada, que tem inverno ameno e bastantes chuvas; Desertos que são caracterizados pela baixa precipitação e Bosques que são facilmente reconhecíveis pelo tipo de folha de suas arvores, que são duras e chamadas de esclerofilas.
Em temperaturas mais altas estão: os desertos mais conhecidos, como (os quentes) no bioma Deserto Subtropical; Savanas, que incluem no Brasil Cerrado e Caatinga, são vegetações com folhas decíduas e que sofrem periódicos incêndios e por fim a Floresta Pluvial Tropical, que inclui Atlântica e Amazônica, que é um bioma de vegetação densa, com muitas chuvas e é aqui que há a maior biovidersidade.
Na imagem abaixo pode-se notar a zonação de biomas terrestres no planeta.

No encontro do continente com oceano há alagados e estuários. Alagados são biomas que misturam água doce com salobra no subsolo, com formação de lagos, mas, sem fluxo superficial de água, como marismas e apicuns. Nos Estuários há esse fluxo superficial de água e consequentemente mais nichos com maior riqueza de espécies.
Os Oceanos constituem um único bioma, que se divide em zonas de acordo com a profundidade, que influenciará na ciclagem de nutrientes e fotossíntese, o que interfere na teia trófica. De 0 a 200 metros de profundidade o litoral compreende: supralitoral, região sempre acima da altamar, ou maré alta; mediolitoral que ocorre na zona entremarés alta e baixa, estando oras coberto oras descoberto e infralitoral, que está sempre coberto pela baixamar, ou maré baixa. A maioria dos seres vivos do oceano ocorrem no litoral. De 200 a 1000 metros está a zona batial e de 1000 a 4000 a abissal, onde gradualmente aumentam pressão, diminui-se temperatura e produtividade. Na zona hadal, abaixo de 4000 metros de profundidade, toda fonte de alimento vem de bactérias quimiotróficas ou de restos de outros animais que submergem e esse ambiente segue até o assoalho marinho.

Fonte: BioInteressante
A afirmação de que a diversidade diminui do equador para os pólos não desqualifica os biomas desses últimos quanto ao conhecimento. Na verdade, pensa-se que isso se deve ao fato do fator climático e estabilidade do meio. Mutações e novos organismos surgem a todo momento, porém, como a estabilidade ambiental é menor no equador há mais espaço para mais nichos serem ocupados e então, novas espécies, porém nos pólos a maioria dos nichos já têm seus organismos bem adaptados e que dominam o local, simplesmente porque o mesmo tem poucas variações. Estudos em ambas as situações são válidos, porque evidenciam e embasam linhas de pesquisa diferentes.
Referências:
RICKLEFS, R.E. A Economia da Natureza. 6 ed. Editora Guanabara Koogan: Rio de Janeiro, 2010.
CH
































{kind=link}